

Índice de la página
1. Structure
2. Formation
3. Function
- Centrosome
- Cilliogenesis
- Asymmetry
- Cellular organization
- Development
Centrioles and basal bodies are structures made up microtubules found in most eukaryote cells. Centrioles, together with pericentriolar material, form centrosomes, and basal bodies form flagella and cilia. Both, centrioles and basal bodies have the same molecular organization and can be interchangeable in the cell. That is, a centriole can move near the plasma membrane and form a cilium, and a basal body can travel to the inner part of the cell and forms a centrosome. Centrioles are involved in the centrosome organization, whereas basal bodies polymerize the microtubules of cilia and flagella. However, centrioles and basal bodies have other secondary associated functions.
1. Structure
In humans, a mature centriole, or basal body, is a cylinder of about 150 to 500 nm height, and about 250 nm in diameter. Thus, centrioles /basal bodies are among the largest molecular structures of the cell. The height is variable between centrioles of different cells and it is yet unknown how it is established. The walls of the cylinder are made up of 9 triplet microtubules. However, there are examples of 9 pair microtubules, as in fly fruit embryos, or even 9 single microtubules, as in C. elegans nematode embryos. In any case, microtubules are always with the same orientation, their plus ends are in one part of the cylinder, and the minus ends in the other. They are referred as distal and proximal ends of the centriole/basal body, respectively, and therefore the cylinder is a polarized structure. In triplet microtubules, only one microtubule, known as microtubule A, has the complete set of 13 protofilaments (rows of tubulin dimers), whereas microtubule B and C has only 10 because they are sharing 3 protofilaments with microtubule A and B, respectively. The distal part of the centriole is formed by the plus ends of microtubules A and B, whereas microtubule C is shorter. In young centrioles, there is a protein structure resembling a cart wheel that provides consistence and spatial organization to 9 triplet microtubules (Figure 1).
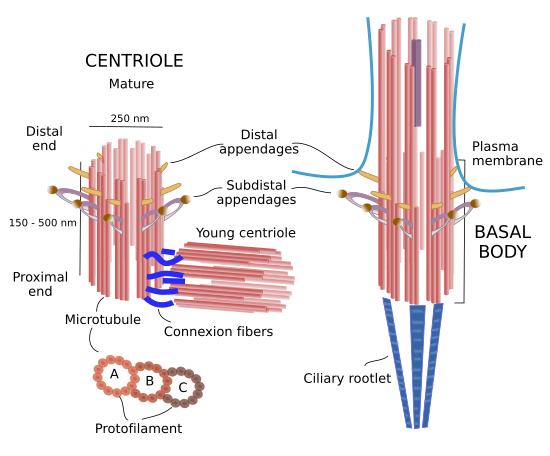
Figure 1. Molecular organization of centrioles / basal bodies.
There are two centrioles in each centrosome: mature and young. Mature centriole bears protein structures known as distal and subdistal appendages. They are found in the distal end of the centriole. Distal appendages are involved with the association of centrioles with the plasma membrane when they migrate to the cell periphery to nucleate cilia and flagella. Subdistal appendages are anchoring sites for microtubules. Basal bodies also have appendages called basal feet and connector fibers, homologous to distal and subdistal appendages. In addition, in the proximal end, basal bodies show long protein rows referred as striated ciliary rootlets. Distal appendages help basal bodies to be anchored to the plasma membrane, whereas striated ciliary rootlets influence the position the cilium or flagellum in the cell periphery.
2. Formation
In the same animal, different number of centrioles and basal bodies can be found in different cell types. Most animal cells contain two centrioles as part of the centrosome, and a basal body as part of a cilium. However, ciliated cells of the trachea epithelium may have hundreds of basal bodies because there are hundreds of cilia. Plant cells, like those of gymnosperms, have lost the capability of synthesizing both centrioles and basal bodies. In some animals it is possible to find cells with and without centrioles. For example, oocyte of mammals and striated skeletal muscle cells lack centrioles.
During cell division, having a proper number of centrioles is essential for most animal cells. If just a pair of centrioles are present during mitosis, there will be a monopolar mitotic spindle and cell division will stop. More than two pairs of centrioles will form multipolar mitotic spindles that produce a wrong segregation of chromosomes (aneuploidy), and it may cause cell malfunctioning or even cell death. Before cell division, all cilia must be deassembled along with their basal bodies. Each of the cell centrioles of the centrosome is used as a platform to nucleate two new centrioles during S phase of the cell cycle. Hence, two pairs of centrioles, and no basal bodies, are found in the cell cytoplasm before mitosis begins. These two pairs of centrioles, along with the pericentriolar matrix, form the two centrosomes that nucleate mitotic spindle microtubules. After cytocinesis, each of the two new cells contain a pair of centrioles, i.e. one centrosome. During G1 and the begining of S phase, there are protein complexes called conexion proteins that keep both centrioles together joined by their proximal ends. The connexion proteins are disorganized after the nucleation of the new two new centrioles, so each pair of centrioles can travel independently through the cytoplasm.
There are two ways of producing new centrioles / basal bodies. The most frequent is forming new centrioles / basal bodies from another one already present, who works as a platform for the nucleation of the new one. For example, after fertilization, sperm usually provides the first centriole for the new animal. Another way to get new centrioles / basal bodies is by nucleation in cells that do not have a previous one. For instance, some cells need to get many basal bodies to form cilia in a short period of time, and mouse embryos may grow until the 64 cell stage without centrioles. In these cases, centrioles / basal bodies nucleate from a non well-known amorphous material called deuterosome.
In centrosomes, the formation of a new centriole, or procentriole, happens near the proximal end (minus ends of microtubules) of the preexisting centriole. A protein cart-wheel like structure is first observed (Figure 2) during S phase of the cell cycle. The cart-wheel has 9 spokes and it needs proteins like Plk4, SAS-6 y STIL. At the end of each spoke, microtubule A is nucleated from a γ-tubulin ring. Microtubules B a C use microtubule A to polymerize, helped by ε- and δ-tubulin. For example, microtubule C is not formed if δ-tubulin is missing. The cart-wheel structure disappears during G1, after mitosis, once the procentrioles get mature. It is unknown how the final length of centrioles is established, but it is clear that only procentrioles can grow in length, which happens during G2 phase of the cell cycle. Procentrioles reach maturity once they polymerize distal and subdistal appendages and reach a proper length.

Figure 2. Growing a new centriole. A procentriole is nucleated from a cart-wheel structure formed near the proximal end of a mature centriole. The cart-wheel structure is observed at the beginning of the S phase of the cell cycle.
Once maturation is fulfilled, centrioles are very stable structures. Centriole stability relies on the lack of tubulin dimers exchange between microtubles and the cytosol because dimers are acetylated and polyglutamilated, and have other modifications, that make them remain as part of the centriole microtubules for very long time. In addition, there are protein connections between triplet microtubules that make more stable all the centriolar structure. Microtubules A and B of basal bodies, however, grow to form the nine microtubule pairs of the cilia / flagela axoneme. The increase in length of these microbutules is due to an internal transport of molecules between the base and tip of the cilium / flagellum.
3. Function
Centrosome
Centrosomes are the main center for organizing and nucleating microtubules in animal cells. Each centrosome is composed of a pair of centrioles and a cloud of surronding proteins known as pericentriolar material. Centriolos are thought important for the organization of centrosomes because they recruit molecules of the pericentriolar material. γ-tubulin rings, which are part of the pericentriolar material, are the real molds to nucleate new microtubules. Both, centrioles and pericentriolar material perform a crucial role during cell division of animal cells by forming the mitotic spindle. However, in some differentiated cells, such as epithelial cells, neurons, and muscle cells, the centrosome is not the main microtubule nucleator. Furthermore, centrosome is not present in most plant cells and yeasts. Actually, the mitotic spindle of plant cells is formed without centrioles.
Ciliogenesis
Cilia are formed from basal bodies by the growing of microtubules A and B of each triplet microtubules. After the cell division is concluded, the older centriole of the pair inherited by the daughter cell migrates to the cell surface and becomes a basal body to nucleate a cilium. In trachea ciliated cells, oviduct and ependymus cells, all of them with hundreds of cilia in their free surfaces, basal bodies are formed independently of centrioles. Actin filaments and microtubules are involved in docking all these new basal bodies to the plasma membrane. Cilia, and therefore basal bodies, must be disorganized before a cell begins cell division, which may prevent basal bodies interfering with the formation of a bipolar mitotic spindle by centrioles.
Cell asymmetry
Asymmetric cell divisions are those where there is an unequal distribution of mother cell components between the two daughter cells. Although centrioles are not completely necessary for cell division, they are needed for asymmetrical cell divisions because they are responsible for the orientation of the mitotic spindle during mitosis. In addition, the daughter cell that bears the pair of centrioles with the older one has a different set of proteins in the pericentriolar material, such as RNA and transcription factors. For example, the daughter cell with the older centriole develops first the cilium and it is able to respond to environmental signals before than the other daughter cell. Thus, it can show a different cell behavior.
Cell organization
The position of centrioles, as part of the centrosome, in the cytoplasm is important for the internal organization of many cell types. It is also important for cell movement because the cell need to have different compartments in the front part compared to the rear part. In astrocytes, the Golgi apparatus is located in the front, whereas the nucleus is found near the rear part of fibroblasts. The centrosome is responsible for the distribution of these two cell compartments.
The place of centrosome, therefore the centrioles, in a particular location of the cytoplasm may be mediated by interactions with microtubules and actin filaments. Usually, there is an interaction of the plus ends of centrosome nucleated microtubules centrosome with the cell periphery, close to the plasma membrane, that influences the position of the centrosome. However, when the centrosome is close to the nucleus, it is the influence of the nuclear envelope what keeps the centrosome in a central position. In some eukaryotic cells, there are fibrous proteins known as striated fibers that physically connect centrioles and nuclear envelope.
Development
Fertilization is the fusion of two cells: one oocyte and one sperm. Only the sperm has centrioles. This centriole first recruits pericentriolar material from the oocyte cytoplasm to form the centrosome. Then, centrosome nucleates and organizes the microtubule system for the movement of the pronuclei (sperm and oocyte nuclei) to find each other and fuse together to form the diploid zygote nucleus. Later, centrosome gets duplicated and the mitotic spindle are built for the first mitotic division. In some species, sperm provides two centrioles, and, curiously enough, in other species like mice there are no centrioles at all after fertilization, neither in the zygote nor in the first blastomeres of the embryo.
Bibliography
Bornens M. 2012. The centrosome in cells and organisms. Science. 335: 422-426.
Gönczy P. 2012. Towards a molecular architecture of centriole assembly. Nature review of molecular and cell biology. 13: 425-535.
Carvalho-Santos Z, Azimzadeh J, Pereira-Leal JB, and Bettencourt-Dias M. 2013. Evolution: Tracing the origins of centrioles, cilia, and flagella. Journal of cell biology. 194: 165-175.
Avidor-Reiss T, Khire A Fishman EL, Kyoung JH. 2015. Atypical centrioles during sexual reproduction. Frontiers in cell and developmental biology. 3:21.
Winey M, O'Toole E. 2015. Centriole structure. Philosophical transactions of the Royal Society of London B biological sciences. 369:1650.