

Muscle tissue produces the body and organs movement. Muscle cells are known as myocytes or muscle fibers. They have the ability to decrease their length, which in turn produces muscle contraction. Muscle cells are organized in bundles or in sheets. The contractile capacity of these cells depends on the association of actin filaments and myosin II motor protein filaments in the cytoplasm.
Muscle cells are divided into three types: skeletal, smooth and cardiac. Skeletal muscle cells are very long cells with dark striations, which are perpendicular to the cell longitudinal axis when observed at light microscopy. Hence, they are also referred as striated skeletal muscle cells. Cardiac muscle cells, or cardiomyocytes, are shorter, branched and also show striations. Smooth muscle cells are fusiform and lack striations, so the name smooth.
1. Striated skeletal muscle

Striated skeletal muscle is also referred as voluntary because it is responsible for conscious or voluntary movements. It is innervated by nerve fibers coming from the central nervous system. Striated skeletal muscle is usually attached to bones either directly or more commonly through tendons. However, there are muscles non connected to bones like those of the eye, upper esophagus and tongue.
Skeletal muscles are made up of striated skeletal muscle cells, also known as muscle fibers or myocytes, as well as connective tissue, nerves and blood vessels. Striated skeletal muscle cells join together to form fascicles, and fascicles form the skeletal muscle (Figures 1 and 2), main responsible for the movements of the animals. Muscle cells are wrapped by a special extracellular matrix known as basal lamina, plus reticular and collagen fibers that together form the endomysium. Each fascicle is surrounded by a layer of connective tissue referred as perimysium, and the whole muscle is covered by the epimysium, which is also connective tissue (Figure 1). Blood vessels and nerve fibers enter the muscle through these connective tissue layers. Nerve fibers rule the muscle contractions.








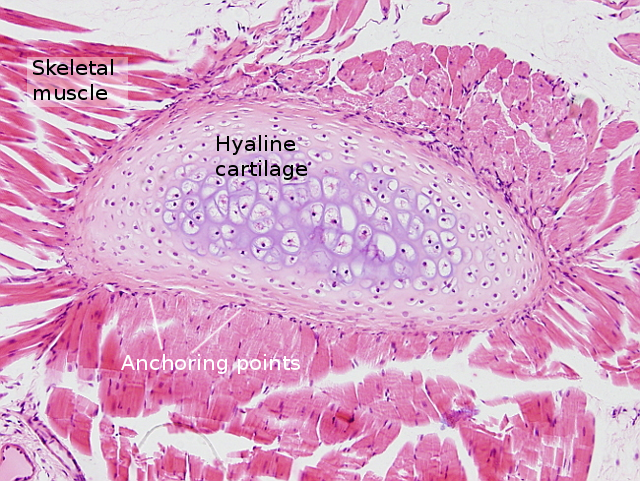
As mentioned above, muscles non-connected to bones, at least non-connected through tendons, show different organization at the anchoring region. Thus, they are attached to the connective tissue of the structures that need to be moved, such as eyes, tongue and esophagus.
Striated skeletal muscle cells are very long and unbranched cells, arranged parallel in sheets and fascicles. They may range from a few to 30 cm in length, and about 10 to 100 µm in diameter. They have many nuclei (syncytium: a cytoplasm shared by two or more nuclei) located just below the plasma membrane. The striations are a consequence of the disposition of actin and myosin filaments in the cytoplasm. Both filaments form bundles known as myofibrils, which are oriented parallel to the long axis of the cell. The overlapping of actin and myosin filaments results in dark bands, and clear bands correspond to just actin filaments.

Although striated skeletal muscle cells can increase and decrease their size (hypertrophy), they can not divide. During postnatal life, the increase in the number of cells (hyperplasia) is consequence of the proliferative activity of satellite cells. These are skeletal muscle stem cells found between the cell membrane of the mature muscle cells and their basal lamina. Satellite cells contain one nucleus, and can divide to give new striated skeletal muscle cells with contractile capacity.
Striated skeletal muscle cells are under the control of neurons (motoneuronas) found in the encephalon and spinal cord. This innervation produces voluntary movements. Each motoneuron can innervate several muscle cells. A motor unit is the group of muscle cells innervated by the axon of a motoneuron, plus the axon itself. Motor units can be large, more than 100 muscle cells innervated by the same axon, or as small as a few tens of innervated muscle cells. The number of neurons of a motor unit depends on the accuracy of the movement that each part of the body needs. More fine movement means smaller units. Furthermore, there are two types of muscle cells regarding the contraction speed. Fast twitch muscle cells are smaller and darker, with a higher concentration of myoglobin and more mitochondria. Slow twitch muscle cells are larger and more clear, and contain less density of mitochondria. The slow twitch muscle cells are active during long-lasting movements and for maintaining the body posture, whereas the fast twitch muscle cells are recruited during short and intense movements. Both types of muscle cells are found in almost every skeletal muscle of the body although with different proportions.
2. Cardiac muscle

Cardiac muscle, or myocardium, is the main component of the heart walls. It pumps the blood through the arteries by contracting the heart ventricle walls.
Cardiac muscle is made up of cardiomyocytes. They show only one nucleus found in a central position, are shorter (about 80 µm) and wider (about 15 µm) than the striated skeletal muscle cells, and are branched cells. Cardiomyocytes also show transversal striations with a pattern similar to that of the striated skeletal muscle cells, with dark strips corresponding to the overlapping of actin and myosin filaments, and light strips being only actin filaments. Cell membrane of striated muscle cells, both cardiac and skeletal, is known as sarcolemma. In mammals, sarcolemma of cardiomyocytes shows many invaginations to form the so-called transverse tubules with a diameter of 5 to 20 nm.
Cardiomyocytes are attached to one another by the so-called intercalated discs, which can be observed as dark strips after general histological staining. Intercalated discs are made up of many cell adhesion complexes, mainly desmosomes and adherent junctions. The role of all these cell adhesion complexes is to keep cardiomyocytes tightly attached between each other, since they are the anchoring sites for cytoskeleton. Gap junctions can also be found in the plasma membrane of adjoining cardiomyocytes, responsible for the synchronization of the heart wall contraction by allowing a direct communication between cytoplasms of neighbor cardiomyocytes. Cardiac muscle is not attached to tendons.
The rhythm of heart beating is under the control of the autonomic nervous system that adjusts the frequency and intensity of the cardiac muscle contractions. However, the rhythmic activity is generated by some special cardiomyocytes working as pacemakers. That is why the cardiac muscle is regarded as a muscle with involuntary contraction. Gap junctions between adjoining muscle cells help to synchronize the contractions by directly connecting their cytoplasms. Because of this, there is no need for nervous innervation of each cardiomyocyte, but just the pacemaker cardiomyocytes. The heart beating frequency is also influenced by hormones.
Cardiomyocytes contain very little glycogen, so they cannot get much energy from glycolysis.Thus, most energy comes from oxidative phosphorylation at mitochondria, which means a very high demand of oxygen. Thus, when there is a cut of the oxygen supply by blood vessel obstruction, cardiomyocytes are quickly damaged.
3. Smooth muscle

The smooth muscle is also known as involuntary muscle. It is found in those organs that do not need voluntary movements such as digestive tract, respiratory system, some glands, gall bladder, urinary bladder, blood and lymphatic vessels, utherus, etcetera.
Smooth muscle cells are 20 to 500 µm in length and 8 to 10 µm in diameter. In the uterine miometrium, they can be as long as 800 µm in length. They are fusiform cells, sometimes with branched ends, and contain one elongated nucleus in the center of the cell. At both poles of the nucleus, there are cytoplasm regions containing most of the cell organelles and some cytoskeleton filaments. The rest of the cytoplasm looks homogeneous and contains the contractile system. Unlike cardiac and skeletal muscles, smooth muscle cells do not organize the actin and myosin filaments in a regular manner, so that there are no striations when observed at light microscopy. The name smooth muscle is because the lack of striations. Smooth muscle cells are wrapped by a layer of extracellular matrix known as basal lamina. There are some parts of the smooth muscle cell without basal lamina that allow direct contact between neighboring cells where gap junctions are established.
In the cytoplasm of smooth muscle cells, there are large dense aggregates of proteins, referred as dense bodies (Figure 3), that contain abundant alpha-actin and are attachment points for actin filaments. The function of dense bodies is similar to Z discs of striated skeletal muscle cells. There are also dense structures in the cell membrane where cytoskeleton filaments are attached. The smaller dark points in the cytoplasm correspond to myosin filaments in transversal view. Scattered actin filaments can be observed in the cytoplasm that together with myosin filaments produce cell contraction. There are around 15 actin filaments per myosin filament (the ratio in the striated skeletal muscle is 6 to 1). Intermediate filaments like desmin and vimentin are also present in the smooth muscle cells. In general, the protein content of smooth muscle cells is 50 % lower than in striated skeletal muscle. The concentration of myosin is particularly lower. However, the concentration of actin and tropomyosin is similar in both types of muscle cells. In the smooth muscle cells, the myosin has to be phosphorylated to activate the actin filaments, so that the contraction speed is slower. Smooth muscle cells do not contain troponin.

She smooth muscle is found in many places of the body, where the smooth muscle cells are organized in different ways according to their functions. Thus, they may be found scattered in some connective tissues, as thin bundles in the dermis attached to the hair follicles, and organized in layers in the digestive duct. The role of smooth muscle in the hollow organs is double: tonic contraction to keep the dimensions of the organ against potentially harmful distensions and contribute to the particular function that organ, such as providing peristaltic movements in the gut and regulation of the blood pressure in the cardiovascular system.
Smooth muscle cell contraction is triggered by axons of the autonomic nervous system. Functionally, smooth muscle cells are organized in two ways: single units and multi-units (Figure 4). In single units, smooth muscle cells are arranged in layers so that the ends of the cells are imbricated among the middle parts of the surrounding cells. Thus, cell membranes of neighbor cells are very close and can form gap junctions. Nerve fibers only make contact with a few cells and the cytoplasm-cytoplasm direct communication synchronizes the contraction of the whole group of connected cells that form the unit. In the multi-unit organization each cell is independently innervated and is isolated from the others by connective tissue.

In some organs like muscle of the digestive duct and trachea, there are intrinsic neuronal plexuses that can independently activate the smooth muscle cells. It happens even with severe spinal cord damages. This neuronal innervation is called intrinsic innervation. However, most smooth muscles also receive inputs from extrinsic inputs from the autonomous nervous system, both sympathetic and parasympathetic. It is remarkable that, whatever the innervation source, there are no specialized structures at the nerve fiber-smooth muscle membrane contacts (unlike the striated skeletal muscle cells, where motor plates are distinct and large structures formed by the incoming axon terminals and the membrane of the cell).
Besides the innervation by nerve terminals, smooth muscle cells may be contracted by autocrine and paracrine chemical signals. There are receptors associated to ionic channels in the plasma membrane for detecting these types of molecules. In addition, there are receptors that sense the cell stretching. Although the smooth muscle may develop a mechanical power similar to striated skeletal muscle cells, the contraction speed is much slower.
Myoepithelial cells found between the basal lamina and the epithelium. They show contractile capacity and help during the release the products synthesized by glands such as salivary, mammary, lachrymal and sweet glands. Myoepithelial cells similar to smooth muscle cells. That is why they are mentioned here. However, myoepithelial cells derive from the ectoderm and the muscle cells develop from mesoderm.
-
Bibliography ↷
-
Bibliography
http://www.uic.edu/classes/phyb/phyb516/smoothmuscleu3.htm
http://advan.physiology.org/content/27/4/201
Berridge MJ. 2012. Cell Signalling Biology. doi:10.1042/csb0001007

-
 Blood
Blood