

El cámbium vascular es un meristemo secundario (y lateral) cuya actividad lleva al crecimiento secundario. Las células del cámbium vascular se dividen formando una banda de células entre el xilema secundario (paredes celulares de color rojo) y el floema secundario (paredes celulares de color verde-azulado). Estas células se dividen periclinalmente, es decir, las dos células hijas se disponen en un plano horizontal, perpendicular a la superficie del órgano (ver figura), originando el floema secundario hacia el exterior y el xilema secundario hacia el interior. Ver la siguiente imagen (Figura 1) del tallo de sambuco a menos aumentos y localizar el cámbium vascular en el tronco.

Se asume que el cámbium vascular está formado por una sola capa de células iniciales, de las cuales derivan tanto xilema como floema secundarios. Hay dos tipos de células iniciales: las fusiformes y las radiales. Las iniciales fusiformes representan más del 90 % de las células del cámbium
Las iniciales fusiformes dan lugar a las células orientadas longitudinalmente en el tronco, denominado sistema axial, que incluyen a las conductoras tanto del floema como del xilema (elementos de los vasos, tubos cribosos y floema, traqueidas y parénquima axial). Dan lugar primero a células que pueden dividirse más veces y que forman la denominada zona cambial. La anchura de la zona cambial indica la tasa de producción de floema y xilema secundarios. Tras la división periclinal de una inicial, cada descendiente se transforma en una célula que se dividirá periclinalmente más veces, produciendo una filas de células que se observan típicamente en las secciones histológicas. Se distinguen bien en gimnospermas, y menos en angiospermas, básicamente porque en estas últimas hay fibras y vasos, incrementando estas últimas mucho su tamaño y distorsionando la fila. Estas células fusiformes iniciales son alargadas con extremos agudos y varían en longitud desde 0.2 mm hasta casi 7 mm. Poseen un núcleo y tienen pared primaria, con numerosos campos de poros primarios.
Las células iniciales radiales se disponen horizontalmente y forman los radios del tronco, células que tendrán misiones como conducción de nutrientes y almacenamiento de sustancias en aquellos troncos que tienen esa capacidad, formando el denominado sistema radial. Cuando se dividen en el cámbium las iniciales radiales, la que queda indiferenciada se puede distinguir porque es menor en diámetro. Las iniciales radiales se forman a partir de las fusiformes iniciales de tres maneras: por divisiones anticlinales de una fusiforme corta, en las fusiformes más largas por múltiples divisiones de una parte de la célula, o por divisiones parciales intermedias. Es importante mantener un equilibrio entre la tasa de iniciales fusiformes y radiales a medida que el cámbium crece en grosor puesto que los radios deben mantenerse en una proporción adecuada respecto a los elementos axiales.
Hay dos maneras de crecer en las células cambiales, y de sus derivados, simplástica e intrusiva. El crecimiento radial es sobre todo simplástico. Durante el crecimiento simplástico las células adyacentes no cambian sus contactos y las células crecen de manera coordinada. El intrusivo es cuando las relaciones de contacto entre células contiguas cambian. El intrusivo es el que más efecto tiene en las propiedades del xilema secundario. Además, de las células cambiales, en el desarrollo del xilema secundario de los árboles hay dos tipos celulares que crecen de manera intrusiva: las fibras y los elementos de los vasos.
Siempre se produce más xilema que floema a partir del cámbium vascular. Ello es debido a que la células progenitoras de xilema sufrirán más divisiones periclinales antes de perder su capacidad mitótica. La tasa de producción de xilema respecto al floema es de 3 a 1 en coníferas, mientras que en dicotiledóneas puede ser de hasta 10 a 1. El xilema secundario está formado sobre todo por células muertas con paredes gruesas compuestas principalmente por celulosa, hemicelulosas y lignina.
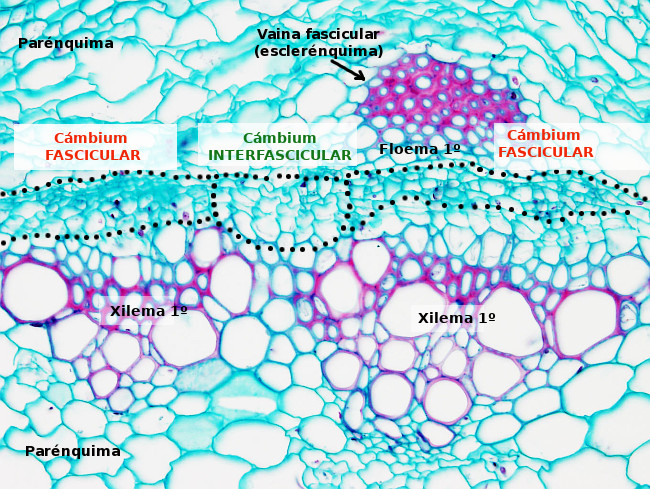
El cámbium vascular se forma por diferenciación del procámbium de los haces vasculares que formará el cámbium fascicular, y por la formación del cámbium interfascicular formado a partir de células parenquimáticas y en algunos casos de la endodermis y colénquima (Figuras 2 y 3). En las raíz se forma a partir del procámbium y del periclo que se encuentra entre las costillas del xilema. La transición entre procámbium y cámbium fascicular en los haces vasculares ocurre tras un cambio en la orientación de los planos de división de las células. Cuando se unen el cámbiun fascicular y el interfascicular se forma el cámbium vascular, el cual se dispone en forma de anillo completo o en forma de bandas paralelas al eje principal de la planta. En órganos maduros no se puede distinguir qué parte del cámbium vascular proviene del cámbium fascicular o del interfascicular sino que el cámbium vascular es un solo anillo continuo de células.

La auxina es una hormona esencial para la formación del cámbium vascular, sobre todo la que se emite desde el ápice del tallo y circula hacia la raíz (basipétala). Si se corta el tallo, el cámbium vascular no se desarrolla. Además la acumulación de auxina entre los haces vasculares favorece la formación del cámbium interfascicular. También es importante para mantener la identidad de cámbium, es decir, para que se mantengan como células indiferenciadas. La mayor concentración de auxina se produce en el cámbium y decrece hacia el floema y el xilema. Hay otras hormonas, como las giberelinas, ácido abcísico y las citocininas, que afectan la actividad de las células cambiales.
Las células cambiales entran en dormancia en especies situadas en regiones con cambios estacionales importantes. El comienzo de esta etapa supone un incremento en grosor de la pared celular, sobre todo de las paredes radiales. La vacuola central se descompone en otras pequeñas, cesa la ciclosis y comienza la síntesis de gránulos de almidón y esferosomas (agregados de lípidos). Los gránulos de almidón son importantes no sólo como reserva energética para el reinicio de la actividad en la próxima estación, sino también como crioprotector. También aumenta el número de mitocondrias, disminuye el retículo endoplasmático, aumentan los ribosomas libres, etcétera.
Durante el periodo de reactivación, la división celular se reinicia y las paredes celulares se hacen más finas, hay cambios en la orientación de los microtúbulos de aleatoria a helicoidal (tanto en las que darán al xilema como al floema), se reinicia la ciclosis, se reconstruye la vacuola central, se hidrolizan los depósitos del citoplasma y orgánulos como el retículo endoplasmático y el aparato de Golgi comienzan su actividad.
Las plantas son capaces de adaptar su crecimiento a estímulos mecánicos como el viento o la presión. Estas respuestas a estímulos mecánicos se denominan conjuntamente como tigmomorfogénesis. Algunas plantas responden disminuyendo la actividad del meristemo apical por lo que son plantas más pequeñas y con troncos más gordos, pero otras responden disminuyendo la actividad cambial y son más delgadas y flexibles. El cámbium también es capaz de responder al peso, es decir, a la cantidad de parte aérea que soporta. A mayor peso, mayor actividad cambial. Incluso el cámbium interfacicular puede ser inducido ejerciendo presión desde arriba del tallo.





{kind=link}