

The shoot grows in length and gives rise to the lateral organs (branches, leaves, flowers, fruits, and seeds). At the same time, it also grows in thickness. Getting thicker may be accomplished with primary and secondary growth. Stems undergoing primary growth are those where the increase in length and thickness depends on the apical shoot meristem and intercalary meristem activities. Stems showing secondary growth increase in diameter by the activity of the vascular cambium and cork cambium. All plant species perform primary growth. Most monocots only show primary growth, whereas many dicots and all gymnosperms have primary and secondary growth.
The activity of the apical shoot meristem is restricted to the tip of the stem. At a short distance, the cells stop proliferating, and the growth occurs by increasing the cell size. This is typical primary growth. However, palms and other monocots may have very thick stems by adding new vascular bundles in distant regions from the stem apical tip and by increasing the number of parenchyma cells. This type of growth is referred to as anomalous growth. It is a consequence of the secondary growth meristem (it is not the vascular cambium), which is found to be more superficial than the vascular bundles. It gives parenchyma cells outward and parenchyma cells and vascular bundles inward. Actually, it is a non-well-delimited region found in the periphery of the stem, containing strands of provascular tissue that give rise to vascular bundles and leaf rays.
The axillary buds are found at the insertion point of the leaf petioles with the stem. They are meristems that remain latent until they develop into branches or flowers. This activation happens when the apical dominance fades out. A combined mix of inhibitory substances is released by the apical caulinar meristem, including hormones (auxin, CKs and SLs), carbohydrates, and other signaling molecules. As the stem elongates, the older axillary buds get far from the apical caulinar meristem influence, and the concentration of inhibitors is progressively lower, which leads to their activation.
Commonly, a stem undergoing primary growth shows the following tissues:
Epidermis. It is usually a single-cell-thick layer covering the stem. Epidermal cells show cutin and suberin in the free cell wall. It is common to have stomata and trichomes, although in a lower number than in the leaves.
Cortex. It is usually a thick layer of parenchyma cells found below the epidermis. Cells can do photosynthesis or store substances. Generally, the outer region of the cortex contains support tissue like sclerenchyma or collenchyma, as in monocots. Occasionally, sclereids, glandular cells, and laticifers can be found in the cortex of some species.
In the cortex, the closest region to the epidermis is known as the hypodermis, which usually has supporting functions. The parenchyma region near the vascular bundles sometimes forms a layer known as the endodermis that shows different features compared with other regions of the cortex. In some non-angiosperm species, a Casparian stripe is also observed in the endodermis. Endodermis cells have been suggested to be involved in the negative gravitropism of the stem, thanks to their content in amyloplasts. That is why this layer of cells is also called the layer of starch. They have also been related to phototropism, the stimulation of the vascular cambium activity, and the elongation of the vascular cells. If a well-developed endodermis is present, an inner pericycle is often observed. Endodermis and pericycle are frequent in aquatic plants.

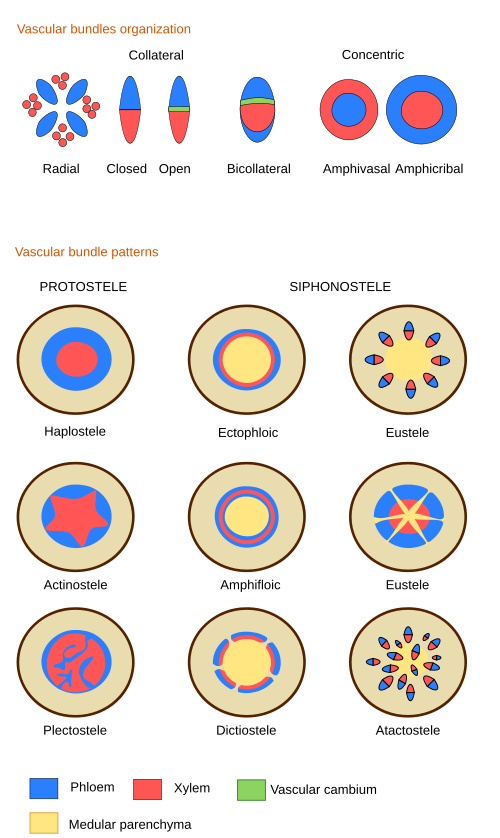
Vascular bundles. Shoots with primary growth may have vascular bundles showing different organizations of the primary vascular tissues: the primary xylem and the primary phloem. This tissues form collateral bundles where the primary phloem is inner and the primary xylem localizes outward. In a typical monocot plant, vascular bundles are scattered through the parenchyma tissue, an organization known as atactosele (ver figura). In typical dicots and gymnosperms, vascular bundles are lined up in a circle, leaving interfascicular parenchyma between each other. This vascular organization is known as the eustele . Only some dicots and conifers form a continuous circle (that is, like a cylinder) of vascular bundles during primary growth. In these species, the xylem is inward and the phloem is outward.
{kind=link}
The mechanical strength of the monocot stems relies on the sclerenchyma fibers found in the vascular bundles and near the epidermis.

During primary growth, the organization of stem and root vascular bundles is different (Figure 1). In the root, xylem and phloem alternate, and metaphloem is inner to protophloem. In the shoot, phloem is outer to xylem, and protoxylem is inner to metaxylem. That is, phloem and xylem change positions, and xylem makes a twist. However, root and shoot vascular bundles have to be connected. It happens at the transition region between the root and the shoot.

In shoots under primary growth and those starting secondary growth, there are vascular bundles directed to the leaves, entering through the petiole. They are known as foliar traces or leaf traces (Figure 2). In this way, continuity is established between the central shoot vascular bundles and those irrigating the leaves, so that leaves are able to get water and minerals and evacuate photosynthesized products. Foliar traces are made up of primary phloem and primary xylem. One leaf may be connected to one or several leaf traces. For example, the leaves of many dicots are connected through 3 to 5 foliar traces.

In species with siphonostele, both ectophloic and amphiphloic (see figure), and in ferns, the vascular cylinder is interrupted above the leaf trace. This discontinuity is known as a leaf gap. At this point, the parenchyma of the cortex and medulla are continuous. If many leaves are in the same shoot, there are many leaf gaps, and then the vascular organization is known as dictiostele.
In eusteles (seed plants under primary growth), there are no leaf gaps because the vascular bundles branch into a leaf trace, and the main vascular bundle is not interrupted. Leaf traces that end up in the same leaf may come from the same vascular bundle (they are known as open systems, typically in helical-arranged leaves). In other species, several leaf traces from different vascular bundles join to form the final leaf trace that enters the leaf.
The vascular bundles in monocots do not run completely parallel to the main axis of the stem, but they grow toward the surface and then get inner again, in a helical way. They send the leaf trace when they are closer to the stem surface. Only some monocots show interrupted vascular bundles by leaf traces. In these cases, the vascular bundle bends toward the stem surface, and it is transformed into a leaf trace.
In plants showing secondary growth, secondary vascular tissue is absent in places where there was a leaf trace. These interruptions are known as leaf gaps. After some time of growing, they are not visible or can only be seen in the inner part of the stem. Once secondary growth is started, no new leaf traces will be formed because no leaves are generated from the vascular bundles in secondary stems.
Medullary region. It is the inner part of the stem. It can be empty (without tissues) or made up of parenchyma tissue.
-
Bibliography ↷
-
Bibliography
Tasaka M, Kato T, Fukaki H. 1999. The endodermis and shoot gravitropism. Trends in plant sciences. 4: 103-107
-