

El tallo crece en longitud formando al mismo tiempo los órganos laterales (ramas, hojas, flores y frutos), y, además, crece en grosor. Este crecimiento en grosor puede ser de dos tipos: crecimiento primario o secundario. Los tallos con crecimiento primario son aquellos en los que el crecimiento en longitud y en grosor depende de los meristemos apicales y de los meristemos intercalares. Mientras que en los tallos con crecimiento secundario, el crecimiento en grosor se debe a los meristemos cámbium vascular y felógeno. Todas las plantas presentan crecimiento primario. La mayoría de las monocotiledóneas sólo tienen este tipo de crecimiento, mientras que la mayoría de las dicotiledóneas y todas las gimnospermas tienen tallos que pasan de un crecimiento primario a otro secundario.
La actividad del meristemo apical ocurre en el ápice de los tallos. A corta distancia del ápice el crecimiento ya no es por proliferación sino por incremento del tamaño de las células. Este es el crecimiento primario típico. Sin embargo, las palmeras y otras monocotiledóneas arbóreas pueden tener tallos muy gruesos debido a un incremento en el número de células parenquimáticas y adición de nuevos haces vasculares en zonas alejadas del ápice del tallo. Este tipo de crecimiento se denomina a veces crecimiento anómalo. Se produce gracias a un meristemo especial que se encuentra externamente a los haces vasculares denominado meristemo de engrosamiento secundario. Este meristemo da hacia fuera células parenquimáticas y hacia dentro células parenquimáticas y células vasculares. Es en realidad una zona un poco difusa localizada en la periferia del tronco. Este meristemo está surcado por hebras de tejido provascular que dará lugar a los haces vasculares y a los radios foliares.
En la unión del peciolo de las hojas con el tronco se encuentran las yemas axilares. Son meristemos que permanecen en estado durmiente hasta que se activan y se convierten en inflorescencias o en ramas. Su activación se produce cuando la dominancia apical disminuye. La dominancia apical es la inhibición que produce el meristemo apical del tallo sobre las yemas axilares próximas. Esta inhición está mediada por un efecto combinado de hormonas (auxinas, CKs y SLs), azúcares y otras moléculas señalizadoras. A medida que el tallo crece y el meristemo apical se aleja de las yemas axilares más antiguas, la concentración de estas moléculas disminuye y la yema se activa.
En general, en un tallo que está en crecimiento primario observamos desde su superficie hasta su eje central los siguientes tejidos:
Epidermis. Es una lámina formada habitualmente por una sola capa de células que presentan cutina y ceras en sus paredes. Es común encontrarse estomas y tricomas, pero generalmente en menor número que en las hojas. Aparte de su papel protector frente a daños mecánicos y frente a la desecación, la epidermis juega un papel fundamental en la cohesión estructural del tallo. Las células epidermis están fuertemente cohesionadas entre sí y soporta presión desde los tejidos internos. La epidermis es responsables de mantener los tejidos internos en su sitio.
Córtex. Es una capa ancha de varias células que se dispone inmediatamente debajo de la epidermis. Sus células son sobre todo parenquimáticas, que pueden ser fotosintéticas o de almacenamiento. Generalmente la región más externa de la corteza lo ocupa un tejido de sostén que puede ser colénquima o esclerénquima, como en el caso de monocotiledóneas. Ocasionalmente en la corteza encontramos esclereidas, células secretoras, y en unos pocos grupos de plantas aparecen estructuras especializadas como los laticíferos.
En la zona parenquimática del córtex, la capa de células que está más próxima a la epidermis se denomina hipodermis, y suele tener función de soporte. La capa de parénquima más próxima a los haces vasculares, generalmente bordeando su parte más externa, tiene unas propiedades diferentes al resto de las de las células parenquimáticas del córtex. A esta capa se le denomina endodermis. Se ha encontrado en algunas especies una banda de Caspary en esta endodermis. A estas células se les ha relacionada con el gravitropismo negativo del tallo (gracias a sus amiloplastos y por ello a esta capa también se le llama capa de almidón), con el fototropismo y con la actividad del cámbium vascular y elongación de las células vasculares. Si hay endodermis suele también haber una capa interior de periciclo. Estas capas son frecuentes en los tallos de plantas acuáticas.

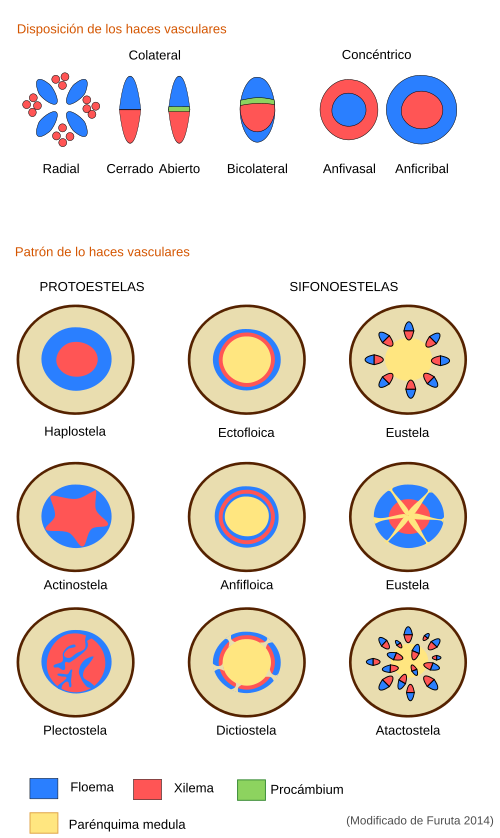
Tejidos vasculares. Los tallos en crecimiento primario de los diferentes grupos de plantas se diferencian entre sí por la disposición de los tejidos vasculares: el xilema y el floema primarios. Éstos se agrupan formando haces colaterales, es decir, el floema primario es externo en relación al xilema. Así, en una monocotiledónea típica los haces se disponen aislados y dispersos en el parénquima, organización denominada atactostela (ver figura), mientras que en las dicotiledóneas y gimnospermas el floema y el xilema primarios se encuentran formando un círculo, dejando entre ellos un parénquima denominado interfascicular, organización denominada eustela. Sólo en algunas plantas dicotiledóneas y coníferas los haces vasculares forman un cilindro completo durante el crecimiento primario, dejando hacia afuera el córtex y hacia dentro la médula. En estos casos el xilema es interno y el floema externo.
{kind=link}


La resistencia mecánica de los tallos de las plantas monocotiledóneas está basada sobre todo en las fibras de esclerénquima de los haces vasculares y de aquellas que se encuentran en las proximidades de la epidermis.
La organización de los haces vasculares es diferente en el tallo primario respecto a la raíz primaria (Figura 1). En la raíz primaria el xilema y el floema se disponen de manera alterna, y dentro del floema, el metafloema es interno respecto al protofloema. En el tallo primaio el floema es externo respecto al xilema, y dentro del xilema, el protoxilema es interno respecto al metaxilema. Es decir, el floema y el xilema cambian sus posiciones y el xilema sufre una rotación. Sin embargo, los haces vasculares de ambas organizaciones, la de la raíz y la del tallo, han de estar conectados, cosa que ocurre en la zona de transición entre raíz y tallo.

En los tallos primarios, y secundarios incipientes, hay haces vasculares que se dirigen hacia las hojas y entran por el peciolo. A estos haces se les llama rastros foliares (Figura 2). Se establece así una continuidad entre los haces vasculares de la estela y los que irrigan las hojas, de modo que las hojas reciben agua y sales minerales y son capaces de evacuar sus productos fotosintéticos. Los rastros foliares tienen floema y xilema primarios. Una hoja puede estar conectada a uno o a varios rastros foliares. Las hojas de muchas dicotiledóneas están comunicadas por tres a cinco rastros foliares.

En plantas con sifonoestelas ectofloicas y anfifloicas (ver figura), y en helechos, el cilindro vascular es discontinuo por encima del rastro foliar. Esta discontinuidad se llama interrupción foliar (en inglés gap). En este punto el parénquima medular y cortical son continuos. Cuando hay muchas hojas en el mismo tallo hay muchas discontinuidades en el cilindro vascular, y entonces a la estela se le denomina dictioestela.
En las eustelas (en plantas con semillas en crecimiento primario) no hay discontinuidades ni interrupciones foliares, es decir, se produce una ramificación del haz vascular para producir el rastro foliar sin que el propio haz quede interrumpido. Los rastros foliares que van a una hoja pueden proceder del mismo haz (se denominan sistemas abiertos, típicos de plantas con hojas dispuestas helicoidalmente) o puede ser el resultado de la fusión de rastros foliares procedentes de diferentes haces. En las atactostelas de monocotiledóneas las hojas suelen estar nutridas por numerosos rastros foliares, que salen de los haces vasculares.
Los haces vasculares de las monocotiledóneas no son totalmente paralelos al eje del tallo sino que se acercan y se alejan de la superficie de una forma helicoidal. Cuando son más periféricos es cuando emite el rastro foliar. Sólo algunas monocotiledóneas tienen haces interrumpidos por las trazas foliares. En estos casos el haz se inclina hacia la superficie del tallo y se transforma en rastro foliar.
En las plantas con crecimiento secundario, allí donde había un rastro foliar no suele haber haces vasculares secundarios. A esto es a lo que realmente que se llama lagunas foliares. A medida que crece el tallo, estas lagunas desaparecen. Luego son estructuras que sólo son visibles en el interior del tallo en crecimiento secundario, normalmente causadas durante el primer año de crecimiento secundario. En los tallos con crecimiento secundario la hojas desaparecen y por tanto ya no habrá rastros foliares.
Médula. Es una región que ocupa la parte más interna del tallo, y puede estar hueca (sin tejidos) u ocupada por tejido parenquimático.
-
Bibliografía ↷
-
Tasaka M, Kato T, Fukaki H. 1999. The endodermis and shoot gravitropism. Trends in plant sciences. 4: 103-107
-